





基因載體是基因工程的核心,也是基因治療中強(qiáng)有力的生物工具,我們先來認(rèn)識(shí)和閱讀載體圖譜吧。
一、載體分類及載體組成元件
載體分類
1、按屬性分類:病毒載體和非病毒載體
病毒載體是一種常見的分子生物學(xué)工具,可將遺傳物質(zhì)帶入細(xì)胞,原理是利用病毒具有傳送其基因組進(jìn)入目的細(xì)胞,進(jìn)行感染的分子機(jī)制。可發(fā)生于完整活體或是細(xì)胞培養(yǎng)中。可應(yīng)用于基礎(chǔ)研究、基因療法或疫苗。用于基因治療和疫苗的病毒載體應(yīng)具備以下基本條件:
(1)攜帶外源基因并能包裝成病毒顆粒;
(2)介導(dǎo)外源基因的轉(zhuǎn)移和表達(dá);
(3)對(duì)人體不致病;
(4)在環(huán)境中不會(huì)引起增殖和傳播。
非病毒載體一般是指質(zhì)粒DNA。
2、按進(jìn)入受體細(xì)胞的類型分類:原核載體、真核載體、穿梭載體(含原核和真核2個(gè)復(fù)制子,能在原核和真核細(xì)胞中復(fù)制,并可以在真核細(xì)胞中有效表達(dá))。
3、按功能分類:克隆載體、表達(dá)載體
克隆載體:具有克隆載體的基本元件(Ori,Ampr,MCS等),可以攜帶DNA-片段或外源基因進(jìn)入受體細(xì)胞并克隆和大量擴(kuò)增DNA-片段(外源基因)的載體。
表達(dá)載體:克隆載體中加入一些與表達(dá)調(diào)控(具有轉(zhuǎn)錄/翻譯所必需的DNA順序)有關(guān)的元件即成為表達(dá)載體。
載體組成元件
1、復(fù)制起始位點(diǎn)Ori:即控制復(fù)制起始的位點(diǎn)。Ori的箭頭指復(fù)制方向,其他元件標(biāo)注的箭頭多指轉(zhuǎn)錄方向(正向)。
2、抗生素抗性基因:可以便于加以檢測(cè),如Amp+ ,Kan+
(1)Ampr:水解β-內(nèi)酰胺環(huán),解除氨芐的毒性。
(2)tetr :可以阻止四環(huán)素進(jìn)入細(xì)胞。
(3)camr:生成氯霉素羥乙酰基衍生物,使之失去毒性。
(4)neor(kanr):氨基糖苷磷酸轉(zhuǎn)移酶,使G418(卡那霉素衍生物)失活。
(5)hygr:使潮霉素β失活。
3、多克隆位點(diǎn):MCS克隆攜帶外源基因片段,它具有多個(gè)限制酶的單一切點(diǎn),便于外源基因的插入。如果在這些位點(diǎn)外有外源基因的插入,會(huì)導(dǎo)致某種標(biāo)志基因的失活,便于篩選。決定能不能放目的基因以及如何放置目的基因。還要再看外源DNA插入片段大小。質(zhì)粒一般只能容納小于10kb的外源DNA-片段。一般來說,外源DNA-片段越長(zhǎng),越難插入,越不穩(wěn)定,轉(zhuǎn)化效率越低。
4、P/E:?jiǎn)?dòng)子/增強(qiáng)子
5、Terms:終止信號(hào)
6、加poly(A)信號(hào):可以起到穩(wěn)定mRNA作用
示例閱讀載體:
pENTER載體
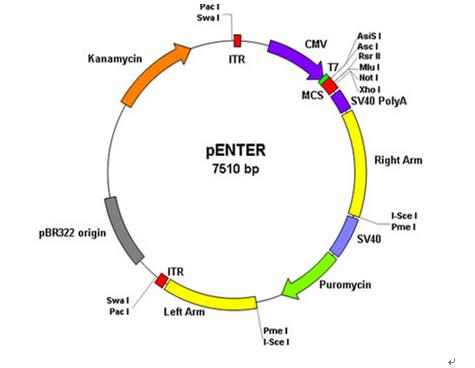
1)human ORF + pENTER載體
2) CMV啟動(dòng)子,T7啟動(dòng)子
3) ORF的C端融合了Flag和His tag
4) 多克隆位點(diǎn),常用AsisI 和 MluI(人源基因上不常見的)
4) SV40 poly(A)加尾信號(hào)
5) SV40啟動(dòng)子啟動(dòng)的puro標(biāo)記
6) 2個(gè)腺病毒ITR序列和2個(gè)與腺病毒Ad5同源的序列,可用于將ORF序列同源重組到腺病毒載體,直接用于腺病毒的生產(chǎn)。
慢病毒載體
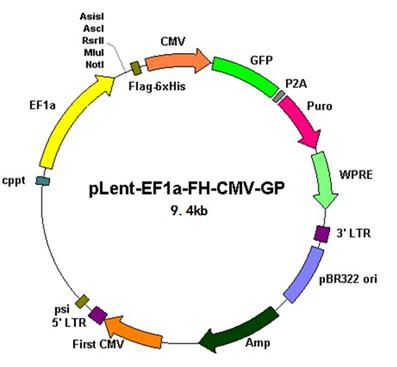
1)EF1a啟動(dòng)目的基因(可添加小標(biāo)簽FLAG或HIS等小標(biāo)簽)
2)CMV啟動(dòng)GFP,非融合抗性篩選標(biāo)記Puro(便于做細(xì)胞株的穩(wěn)篩)
3)WPRE(來自土撥鼠肝炎病毒的轉(zhuǎn)錄后調(diào)節(jié)元件),放置在目的基因的上游,能加強(qiáng)轉(zhuǎn)基因的表達(dá)
4) cPPT(來自HIV-1整合酶基因),增加慢病毒在宿主基因組的拷貝數(shù),提高病毒滴度
5)第三代慢病毒載體,載體中還包含HIV-1基因組中的順式調(diào)節(jié)元件(ψ 包裝 信號(hào)和LTR 長(zhǎng)末端重復(fù)序列,其中3’LTR增強(qiáng)子功能缺失,5’LTR中U3區(qū)替代為CMV,更加安全的病毒載體)
腺相關(guān)病毒載體
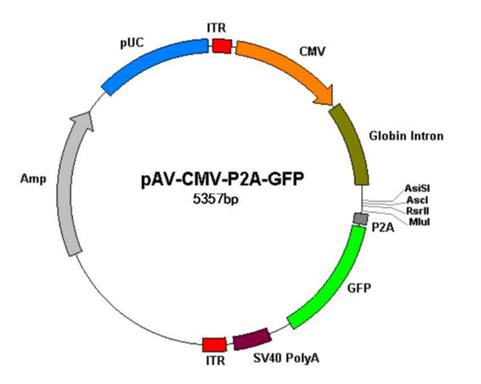
AAV表達(dá)載體包括了中兩個(gè)ITR + CMV(可替換其他組織特異性啟動(dòng)子,見改造載體部分)+ globin intron 內(nèi)含子序列 + 目的基因 + poly A序列
二、選擇和準(zhǔn)備載體
選擇載體主要依據(jù)構(gòu)建的目的,同時(shí)還要考慮載體中應(yīng)有合適的限制酶切位點(diǎn)等。如果構(gòu)建的目的是要表達(dá)一個(gè)特定的基因,則要選擇合適的表達(dá)載體。選用哪種載體,還是要結(jié)合目的基因及載體特點(diǎn)以實(shí)驗(yàn)?zāi)康臑闇?zhǔn)繩。
載體選擇主要考慮下述3點(diǎn):
1、構(gòu)建DNA重組體的目的,克隆擴(kuò)增/表達(dá)表達(dá),選擇合適的克隆載體/表達(dá)載體。
2、載體的類型:
(1)克隆載體的克隆能力—據(jù)克隆片段大小(大選大,小選小)。尤其對(duì)于病毒載體來說,載體包裝容量的大小是評(píng)估能否成功包裝病毒的首要因素。
①腺病毒可以插入長(zhǎng)約8kb的外源基因。目前,我們包裝過長(zhǎng)度為7.5kb外源基因的腺病毒。
②慢病毒的包裝容量約為6kb(包含載體帶有的其他抗性基因或報(bào)告基因),但是2kb以上的外源基因包裝慢病毒難度就增加了,增大1kb會(huì)下降1個(gè)單位數(shù)量級(jí)的滴度,故2kb以上的基因包裝慢病毒需要進(jìn)行評(píng)估。此外,毒性蛋白、凋亡蛋白和膜蛋白(作為受體、轉(zhuǎn)運(yùn)蛋白行使功能)等由于其本身的功能,包裝可能會(huì)有一定難度。
③AAV的總包裝容量是4.7 kb(包含載體中兩個(gè)ITR約0.3kb +具體啟動(dòng)子 + 內(nèi)含子約0.6kb + 具體熒光標(biāo)簽 + polyA約0.2kb),所以插入目的基因一般約2kb 左右。由于AAV包裝容量有限,目的基因大于2kb,可考慮去除內(nèi)含子,因?yàn)閮?nèi)含子只是有助于插入的目的基因穩(wěn)定表達(dá)。
(2)確定表達(dá)蛋白的目的,然后再來選擇優(yōu)勢(shì)的表達(dá)質(zhì)粒。一般分為原核表達(dá)和真核表達(dá)質(zhì)粒,前者主要用于體外純化蛋白,如帶His-tag的PET系列,帶GST-Tag的PGEX系列,選用不同的表達(dá)載體,就得建立相應(yīng)的純化系統(tǒng),包括緩沖液的配置、珠子/柱子的選擇等等,還得考慮目的蛋白的可溶性,一般大tag的融合蛋白可溶性要好一些,如GST的。這種原核純化的蛋白一般用來制備單一的、需要量大的蛋白。真核表達(dá)載體一般是細(xì)胞、體內(nèi)表達(dá)等需要時(shí)所用,只需要將它放到細(xì)胞或體內(nèi)細(xì)胞即可,唯1考慮的是它的啟動(dòng)子的選擇,常用都是cmv啟動(dòng)子的,表達(dá)效率較高,也有多種啟動(dòng)子經(jīng)過改造的表達(dá)載體,一般用來表達(dá)需要調(diào)控表達(dá)時(shí)間及表達(dá)量的蛋白。
3、載體MCS中的酶切位點(diǎn)數(shù)與組成方向因載體不同而異,適應(yīng)目的基因與載體易于連接,不產(chǎn)生閱讀框架錯(cuò)位。
①選分子量小的質(zhì)粒,即小載體(1-1.5kb)→不易損壞,在細(xì)菌里面拷貝數(shù)也多(也有大載體);
②一般使用松弛型質(zhì)粒在細(xì)菌里擴(kuò)增不受約束,一般10個(gè)以上的拷貝,而嚴(yán)謹(jǐn)型質(zhì)粒<10個(gè);
③必需具備一個(gè)以上的酶切位點(diǎn),有選擇的余地;
④必需有易檢測(cè)的標(biāo)記,多是抗生素的抗性基因。
選擇載體還需注意:
無標(biāo)簽載體,起始/終止密碼子都要添加!
標(biāo)簽在N端的載體,終止密碼子要添加!
標(biāo)簽在C端的載體,起始密碼子要添加!
(詳細(xì)解讀如下)
①對(duì)于無起始密碼子和終止密碼子的基因,插入無標(biāo)簽的載體要加入起始密碼子和終止密碼子。
②載體N端有標(biāo)簽,上游引物要對(duì)讀碼框;下游引物要加終止密碼子(為了保證基因的表達(dá),少翻譯載體序列) 。
鏈接:對(duì)讀碼框是為了防止移碼,是不是移碼要看載體圖譜,看酶切位點(diǎn)。從ATG開始,要保證三個(gè)堿基編碼的氨基酸不能影響插入目的基因的正常編碼,若影響了插入目的基因的正常編碼就要在設(shè)計(jì)引物的時(shí)候在酶切位點(diǎn)前或者后插入一個(gè)或者兩個(gè)堿基,總之是不能影響插入目的基因的正常編碼蛋白。有的酶切位點(diǎn)含有起始密碼子,如果標(biāo)簽在C端像Nco I就有一個(gè)ATG,這時(shí)候需要加堿基在酶切位點(diǎn)后面。
③載體C端有標(biāo)簽,上游引物要加起始密碼子,且考慮是否加Kozak序列(真核表達(dá)系統(tǒng));下游引物要對(duì)讀碼框,并且要去掉基因本身的終止密碼子(保證C端標(biāo)簽表達(dá))。
鏈接:Kozak序列是位于真核生物mRNA 5’端帽子結(jié)構(gòu)后面的一段核酸序列,通常是GCCGCCRCC(常見的序列是GCCACC),位于起始密碼子(ATG)之前。它可以與翻譯起始因子結(jié)合而介導(dǎo)含有5’帽子結(jié)構(gòu)的mRNA翻譯起始。在真核生物中,該序列相對(duì)比較保守。對(duì)應(yīng)于原核生物的SD序列(原核基因在mRNA5’端起始密碼子AUG上游一段對(duì)mRNA的翻譯起作用的序列)。
添加Kozak序列的好處:核糖體需要Kozak序列進(jìn)行穩(wěn)定結(jié)合有助于提高真核基因的轉(zhuǎn)錄和翻譯的效率。
三、改造載體
1、改造啟動(dòng)子:一般在過表達(dá)載體中CMV屬于廣譜型的強(qiáng)啟動(dòng)子,具體可根據(jù)實(shí)驗(yàn)需求,選用不同啟動(dòng)子,尤其是對(duì)于AAV載體構(gòu)建時(shí)選用組織特異性啟動(dòng)子。
啟動(dòng)子名稱 | 啟動(dòng)子大小 | 組織特異性 |
ALB | 2.4kb | 肝臟特異性 |
CAG | 944bp | 廣泛表達(dá)的強(qiáng)啟動(dòng)子 |
CamKIIa | 1.2kb | 大腦皮層和海馬神經(jīng)元特異性啟動(dòng)子 |
CMV | 0.6kb | 廣泛表達(dá)的強(qiáng)啟動(dòng)子 |
EF1A | 1.2kb | 廣泛表達(dá)的啟動(dòng)子(尤其在體內(nèi)穩(wěn)定表達(dá)) |
GFAP | 1.0kb | 星形膠質(zhì)細(xì)胞特異性啟動(dòng)子 |
aMHC | 0.4kb | 心肌α肌球蛋白重鏈啟動(dòng)子 |
cTNT | 702bp | 心肌特異性 |
Synapsin | 471bp | 成熟神經(jīng)元特異性啟動(dòng)子 |
Rpe65 | 700bp | 視網(wǎng)膜特異性 |
3Xenhancer McK | 728bp | 小鼠中肌肉特異性啟動(dòng)子 |
NSE | 1.3kb | 神經(jīng)元特異性烯醇化酶啟動(dòng)子 |
2、實(shí)驗(yàn)?zāi)康牡男枰?/p>
①構(gòu)建過表達(dá)or干擾載體
根據(jù)實(shí)驗(yàn)用途選擇載體,如過表達(dá)載體通常選用CMV啟動(dòng)子,CMV因其集中了細(xì)胞轉(zhuǎn)錄因子結(jié)合位點(diǎn)而具有異常高的活性,是*的真核表達(dá)中的增強(qiáng)子,一般插入某個(gè)基因的CDS區(qū)到CMV啟動(dòng)子下游,CMV負(fù)責(zé)啟動(dòng)該基因的表達(dá),從而達(dá)到調(diào)高該基因表達(dá)的作用;而U6和H1更多的是用于shRNA的啟動(dòng)來達(dá)到敲低一個(gè)基因的作用,U6和H1啟動(dòng)子都是RNA聚合酶Ⅲ依賴的啟動(dòng)子,其特點(diǎn)是啟動(dòng)子自身元素均位于轉(zhuǎn)錄區(qū)的上游,適合于表達(dá)約21個(gè)核苷酸和約50個(gè)核苷酸莖環(huán)結(jié)構(gòu)(stem loop),RNA聚合酶Ⅲ有明確的起始和終止序列,當(dāng)RNA聚合酶Ⅲ到連續(xù)4個(gè)或5個(gè)T時(shí),它指導(dǎo)的轉(zhuǎn)錄就會(huì)停止。
我們公司的shRNA病毒載體(包括腺病毒載體、慢病毒載體和腺相關(guān)病毒載體)是由U6或H1 啟動(dòng)shRNA 實(shí)現(xiàn),并連接了報(bào)告基因,可以實(shí)時(shí)監(jiān)測(cè)載體的轉(zhuǎn)染效率。構(gòu)建干擾載體針對(duì)目的基因(目的基因需要大于0.7kb,若小于0.7kb需要評(píng)估來確保能設(shè)計(jì)出8條shRNA序列),設(shè)計(jì)并提供4個(gè)針對(duì)靶基因的shRNA產(chǎn)品,確保4個(gè)shRNA中的1個(gè)產(chǎn)品在細(xì)胞感染效率達(dá)80%以上的前提下,在mRNA水平至少有70%的沉默效果;也可以按照客戶的要求來設(shè)計(jì),由客戶確定shRNA 序列的長(zhǎng)度,對(duì)于每條選定的靶序列,設(shè)計(jì)正義鏈和反義鏈,以 loop莖環(huán)結(jié)構(gòu)相連,合成shRNA。
②是否攜帶GFP等熒光標(biāo)簽
攜帶熒光標(biāo)簽基本目的是為了觀察感染目的細(xì)胞的效率;攜帶該標(biāo)簽的好處之一是不用破碎組織細(xì)胞和不加任何底物,直接通過熒光顯微鏡就能在活細(xì)胞中發(fā)出綠色熒光,實(shí)時(shí)顯示目的基因的表達(dá)及定位情況,而且熒光性質(zhì)穩(wěn)定。但是,由于GFP標(biāo)簽比較大(約720bp),這種大標(biāo)簽在位置上對(duì)目的基因造成了空間位阻作用,對(duì)蛋白的空間結(jié)構(gòu)產(chǎn)生一定影響,從而不利于目的基因蛋白本身的活性及正常功能的發(fā)揮。此外,還需要結(jié)合蛋白本身的特性,比如該蛋白為具有切割活性的蛋白,即使融合了GFP,標(biāo)簽很有可能會(huì)被破壞,終究會(huì)影響GFP的熒光及基本功能。一般不建議融合這么大的標(biāo)簽,實(shí)驗(yàn)需要帶熒光標(biāo)簽,建議構(gòu)建載體時(shí)通過P2A非融合。
③是否融合6×His、HA、Myc、FLAG等小標(biāo)簽
為了做免疫共沉淀實(shí)驗(yàn)或者WB檢測(cè)蛋白表達(dá),需要加融合小標(biāo)簽。多數(shù)情況下,由于多肽標(biāo)簽相對(duì)較小,對(duì)目的蛋白質(zhì)結(jié)構(gòu)影響小。但是在蛋白合成肽鏈的過程中,如果影響了蛋白的折疊,也會(huì)造成目的蛋白活性的下降甚至功能的喪失。
④是否含有表達(dá)系統(tǒng)元件,即啟動(dòng)子-核糖體結(jié)合位點(diǎn)-克隆位點(diǎn)-轉(zhuǎn)錄終止信號(hào)。
鏈接:?jiǎn)?dòng)子:促進(jìn)DNA轉(zhuǎn)錄的DNA序列,這個(gè)DNA區(qū)域常在基因或操縱子編碼區(qū)的上游,是DNA分子上可以與RNA聚合酶特異性結(jié)合并使之開始轉(zhuǎn)錄的部位,但啟動(dòng)子本身不被轉(zhuǎn)錄。
增強(qiáng)子:為真核基因組(包括真核病毒基因組)中的一種具有增強(qiáng)鄰近基因轉(zhuǎn)錄過程的調(diào)控順序,其作用與其所在的位置或方向無關(guān)。即在所調(diào)控基因上游或下游均可發(fā)揮作用。
沉默子:負(fù)增強(qiáng)子,負(fù)調(diào)控序列。
核糖體結(jié)合位點(diǎn)/SD序列:mRNA的起始AUG上游約8~13核苷酸處,存在一段由4~9個(gè)核苷酸組成的共有序列,可被16SrRNA通過堿基互補(bǔ)精確識(shí)別的序列被稱為核糖體結(jié)合位點(diǎn)/SD序列是對(duì)原核生物而言,在mRNA 5’ 端起始密碼子AUG上游一段對(duì)mRNA的翻譯
起作用的序列。
轉(zhuǎn)錄終止順序(終止子)/翻譯終止密碼子:結(jié)構(gòu)基因的后一個(gè)外顯子中有一個(gè)AATAAA的保守序列,此位點(diǎn)下游有一段GT或T富豐區(qū),這2部分共同構(gòu)成poly(A)加尾信號(hào)。
3、載體中P2A和IRES的選擇:
| IRES和P2A的比較 | |
| IRES | P2A |
定義 | IRES內(nèi)部核糖體進(jìn)入位點(diǎn)序列:是一段約500bp的核酸序列,這類RNA序列能折疊成類似于起始tRNA的結(jié)構(gòu),從而介導(dǎo)核糖體與RNA結(jié)合,能獨(dú)立的起始蛋白翻譯。 | P2A肽(2A peptide)是一種可"自我剪切"的短小肽鏈,初在FMDV中發(fā)現(xiàn),約22個(gè)氨基酸,2A肽可在蛋白翻譯時(shí)通過核糖體跳躍從自身后2個(gè)氨基酸C末端斷裂。 |
優(yōu)點(diǎn) | IRES被放置于兩個(gè)ORF之間的時(shí)候,可以同時(shí)表達(dá)這兩個(gè)ORF。由于兩個(gè)ORF分別有自己的起始密碼子和終止密碼子,所以會(huì)翻譯兩個(gè)獨(dú)立的、沒有經(jīng)過修飾的蛋白。 | 兩個(gè)基因(ORF)通過2A多肽鏈連接成為一個(gè)ORF,mRNA翻譯成一個(gè)融合蛋白,但這兩個(gè)融合蛋白會(huì)被識(shí)別2A的蛋白酶切成兩個(gè)蛋白。這兩個(gè)蛋白的摩爾比理論上是1:1。 |
缺點(diǎn) | IRES的存在有時(shí)候會(huì)影響mRNA的結(jié)構(gòu),同時(shí)由于IRES和mRNA的5’CAP對(duì)核糖體/或翻譯起始復(fù)合物的結(jié)合力不同,IRES后面的ORF翻譯蛋白的水平有可能與IRES前面的ORF蛋白水平不一致。有的時(shí)候前面的ORF表達(dá)很好,但后面的ORF表達(dá)水平不高;有的時(shí)候后面的ORF和/或IRES會(huì)影響前面ORF的表達(dá),甚至前面的ORF根本不表達(dá)。 | 2A是一個(gè)大約22個(gè)氨基酸的多肽。蛋白酶切割會(huì)發(fā)生在2A多肽C端的甘氨酸(G)和脯氨酸(P)之間。所以,前一個(gè)蛋白的尾巴上會(huì)留下一個(gè)20多個(gè)氨基酸的多肽。后面一個(gè)蛋白的N端會(huì)留下一個(gè)多余的脯氨酸。尤其是第一個(gè)蛋白,如果是一個(gè)小分子(比如分泌型的細(xì)胞因子),可能其功能會(huì)受到這20多個(gè)氨基酸的影響。 |
建議 | IRES序列后基因的表達(dá)效率顯著低于IRES序列前基因的表達(dá),所以與IRES相比,2A肽具有以下優(yōu)點(diǎn):(1)2A序列短,能夠有效地實(shí)現(xiàn)連接基因之間的共表達(dá);(2)位于2A下游的基因同樣可以獲得很高的表達(dá)水平。 | |
四、影響載體選擇的因素:
? 原核表達(dá) or 真核表達(dá)
? 細(xì)胞實(shí)驗(yàn) (過表達(dá) or 干擾、瞬時(shí)表達(dá) or穩(wěn)轉(zhuǎn)株、基因大小、熒光、藥篩 )
?動(dòng)物實(shí)驗(yàn) (過表達(dá) or 干擾、注射部位、基因大小、觀察周期 )